
|
Muscle Cell Structure and Physiology | |
 |
The functional characteristics of a skeletal muscle cell:
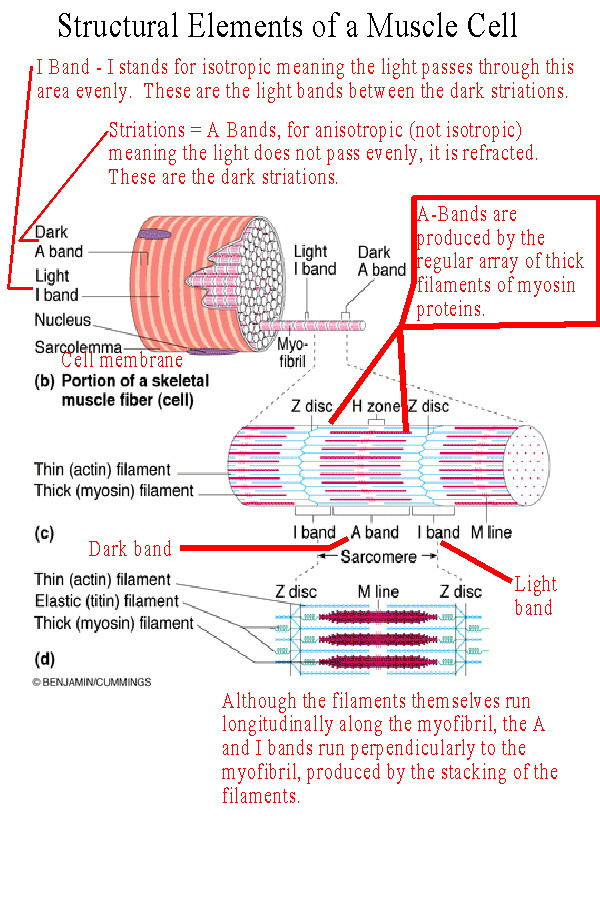
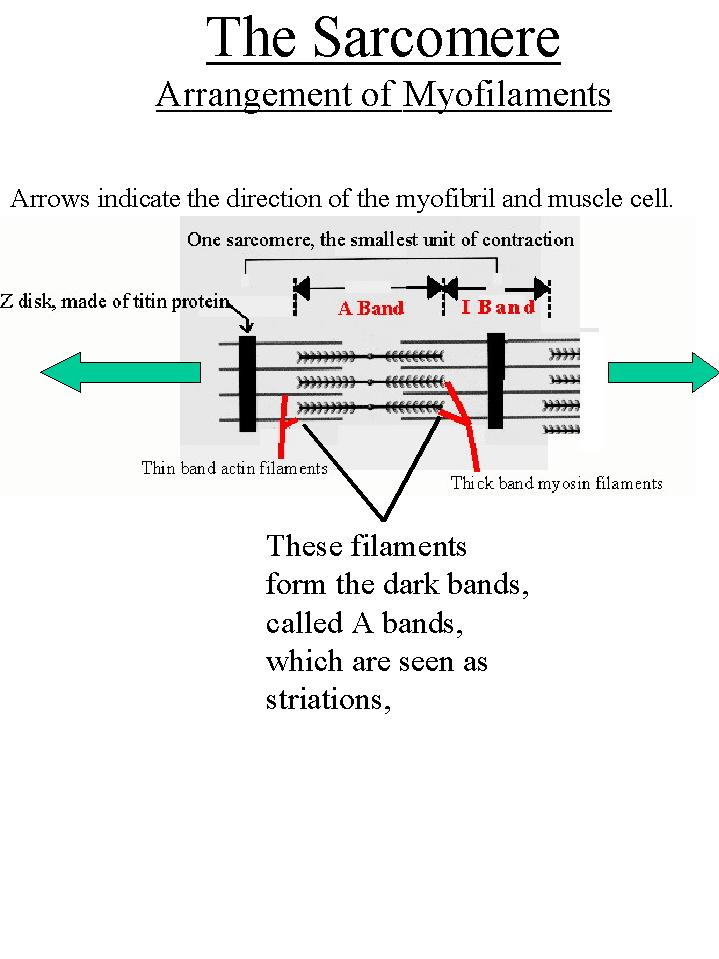
The cell membrane is called the sarcolemma. This membrane is structured to receive and conduct stimuli. The sarcoplasm of the cell is filled with contractile myofibrils and this results in the nuclei and other organelles being relegated to the edge of the cell. Myofibrils are contractile units within the cell which consist of a regular array of protein myofilaments. Each myofilament runs longitudinally with respect to the muscle fiber. There are two types: the thick bands and the thin bands. Thick bands are made of multiple molecules of a protein called myosin. The thin bands are made of multiple molecules of a protein called actin. The thin actin bands are attached to a Z-line or Z-disk of an elastic protein called titin. The titin protein also extends into the myofibril anchoring the other bands in position. From each Z-line to the next is a unit called the sarcomere. The sarcomere is the smallest contractile unit in the myofibril. Sarcomeres contract because the Z-lines move closer together. As the sarcomeres contract the myofibrils contract. As the myofibrils contract the muscle cell contracts. And as the cells contract the entire muscle contracts. The arrangement of the thick myosin filaments across the myofibrils and the cell causes them to refract light and produce a dark band known as the A Band. In between the A bands is a light area where there are no thick myofilaments, only thin actin filaments. These are called the I Bands. The dark bands are the striations seen with the light microscope. |
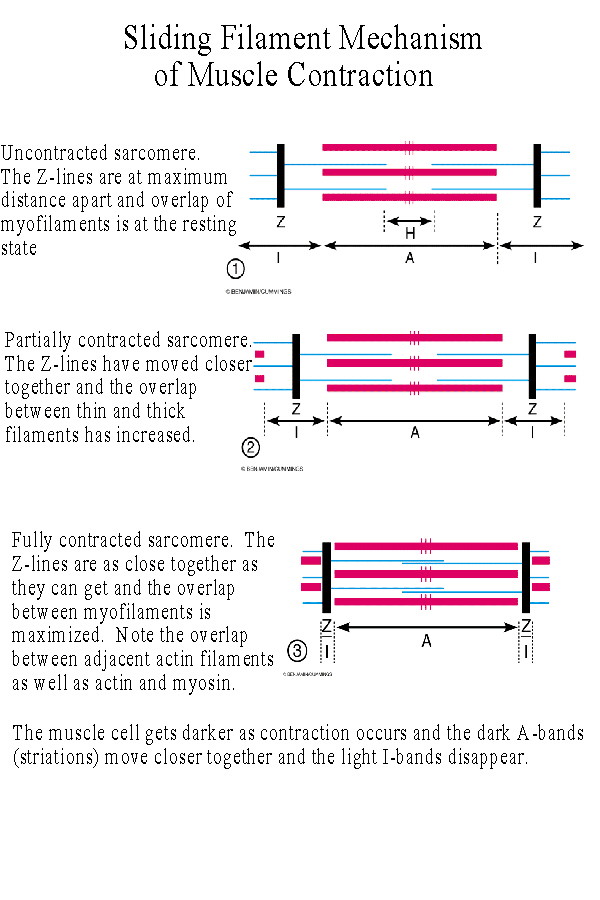 |
When a muscle contracts the light I bands disappear and the dark A bands move closer together. This is due to the sliding of the myofilaments against one another. The Z-lines pull together and the sarcomere shortens as above. |
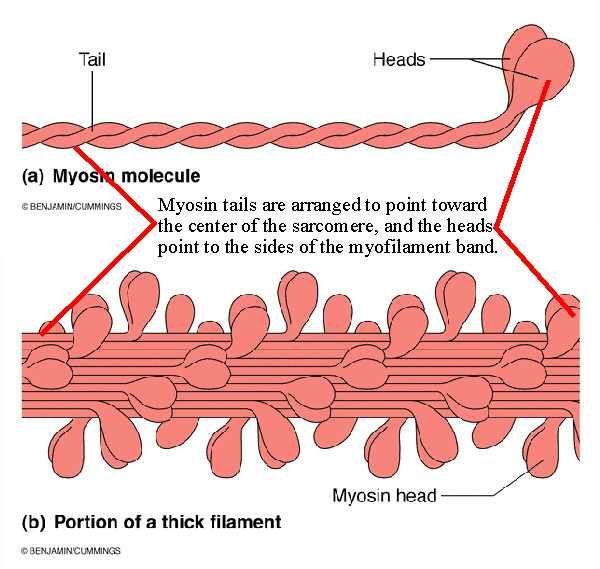 |
The thick myosin bands are not single myosin proteins but are made of multiple
myosin molecules. Each myosin molecule is composed of two parts: the globular
"head" and the elongated "tail". They are arranged to form the thick bands as
shown in Figure 9.3 a and b, modified.
It is the myosin heads which form crossbridges that attach to binding sites on the actin molecules and then swivel to bring the Z-lines together. |
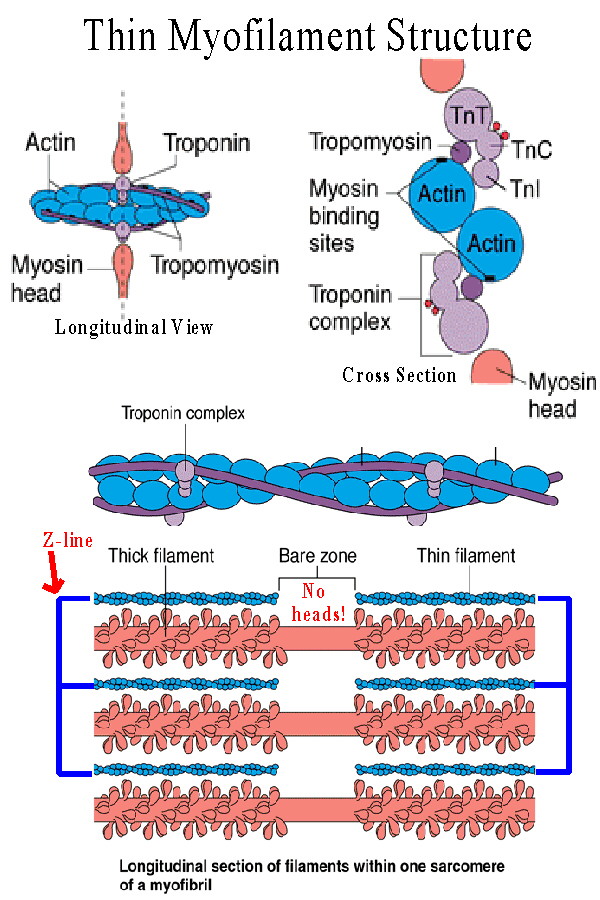 |
Likewise the thin bands are not single actin molecules. Actin is composed of
globular proteins (G actin units) arranged to form a double coil which produces
the thin filament. Each thin actin myofilament is wrapped by a tropomyosin
protein, which in turn is connected to the troponin complex. (See Figures 9.3 c
and 9.7).
The tropomyosin-troponin combination blocks the active sites on the actin molecules preventing crossbridge formation. The troponin complex consists of three components: TnT, the part which attaches to tropomyosin, TnI, an inhibitory portion which attaches to actin, and TnC which binds calcium ions. When excess calcium ions are released they bind to the TnC causing the troponin-tropomyosin complex to move, releasing the blockage on the active sites. As soon as this happens the myosin heads bind to these active sites. |
 |
Events in Muscle Contraction - the sequence of events in crossbridge formation:
(See Figure 9.8, modified at left)
1) In response to Ca+2 release, the troponin-tropomyosin complex removes its block from actin, and the myosin heads immediately bind to active sites. 2) The myosin heads then swivel, the Working Stroke, pulling the Z-lines closer together and shortening the sarcomeres. As this occurs the products of ATP hydrolysis, ADP and Pi, are released. 3) ATP is taken up by the myosin heads as the crossbridges detach. If ATP is unavailable at this point the crossbridges cannot detach and release. Such a condition occurs in rigor mortis, the tensing seen in muscles after death, and in extreme forms of contracture in which muscle metabolism can no longer provide ATP. 4) ATP is hydrolyzed and the energy transferred to the myosin heads as they cock and reset for the next stimulus. |
|
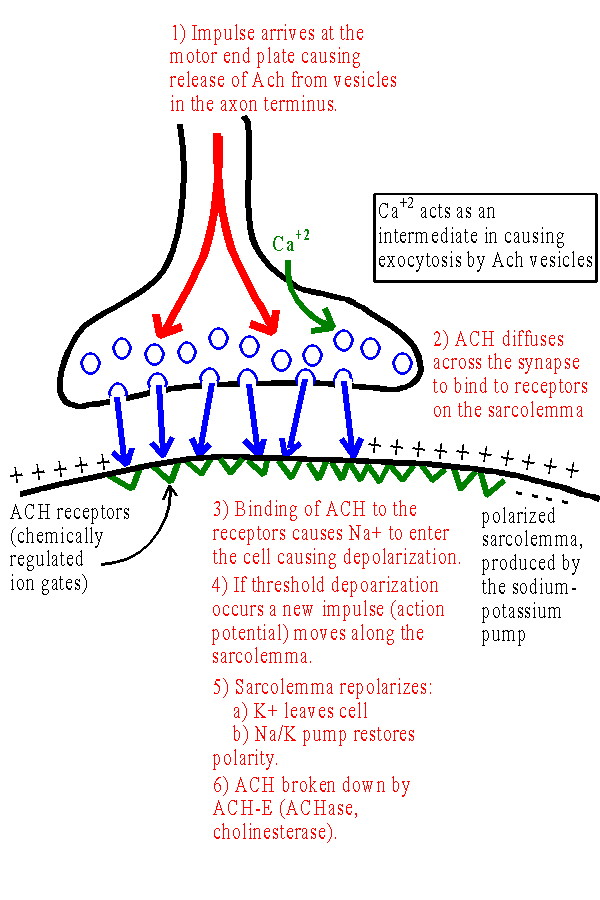
Excitation-Contraction Coupling: the Neuromuscular Junction
Each muscle cell is stimulated by a motor neuron axon. The point where the axon terminus contacts the sarcolemma is at a synapse called the neuromuscular junction. The terminus of the axon at the sarcolemma is called the motor end plate. The sarcolemma is polarized, in part due to the unequal distribution of ions due to the Sodium/Potassium Pump. (See Excitability and Membrane Potential] and Figure 9.9) 1) Impulse arrives at the motor end plate causing release of ACH from vesicles in the axon terminus. Ca+2 acts as an intermediate in causing exocytosis by ACH vesicles into the synapse. 2) ACH diffuses across the synapse to bind to receptors on the sarcolemma. 3) Binding of ACH to the receptors opens chemically regulated ion gates. This causes Na+ to enter the cell producing depolarization. 4) If threshold depolarization occurs, a new impulse (action potential) moves along the sarcolemma. (This occurs because voltage regulated ion gates open as a result of the depolarization - See Figure 9.10) 5) The sarcolemma repolarizes: a) K+ leaves cell (potassium channels open as sodium channels close) returning positive ions to the outside of the sarcolemma. (More K+ actually leaves than necessary and the membrane is hyperpolarized briefly. This causes the relative refractory period) b) Na+/K+ pump restores resting ion distribution and membrane potential. 6) ACH broken down by ACH-E (a.k.a. ACHase, cholinesterase). This permits the receptors to respond to another stimulus. Anti-cholinesterase toxins cause paralysis by leading to blockage of receptors by ACH. |
| Excitation-Contraction Coupling: The action potential and release of calcium.
The action potential is a self-propagated, all-or-none movement of depolarization along the membrane. (See Figure 9.10) All-or-none means that there are not different size action potentials. You either have one or you don't. As the action potential passes along the sarcolemma it causes release of Na+ into the cell by voltage-regulated ion gates, just as the chemically-regulated gates when stimulated by ACH. As the action potential passes along the sarcolemma it enters the T-tubules which occur at each Z-line. (See Figure 9.4) The T-tubules are membranes which run across the cell (T for transverse) connecting to the sarcolemma. The T-tubules allow the action potential to continue into the cell interior. At points along the T tubules they attach to the sarcoplasmic reticulum, a system of membrane channels inside the sarcoplasm. When the action potential moves along the T tubules it causes the sarcoplasmic reticulum to release Ca+2 which is sequestered by the SR. The SR pumps calcium like the sarcolemma pumps sodium and releases it into the sarcoplasm when stimulated by the action potential. This causes the sliding of filaments as outlined above. | |
| Excitation-Contraction Coupling: Summary of Events.
See Figure 9.11 and class diagram. | |
{kind=link}
){kind=link}
){kind=link}
){kind=link}
){kind=link}
{kind=link}