
Click on ![]() for respiratory histology module. Histology linked to
for respiratory histology module. Histology linked to ![]()
Click on  for pathology images related to discussion.
for pathology images related to discussion.
|
The Respiratory System | |
|
Respiration involves several components: Ventilation - the exchange of respiratory gases (O2 and CO2) between the atmosphere and the lungs. This involves gas pressures and muscle contractions. External respiration - the exchange of gases between the lungs and the blood. This involves partial pressures of gases, diffusion, and the chemical reactions involved in transport of O2and CO2. Internal respiration - the exchange of gases between the blood and the systemic tissues. This involves the same processes as external respiration. Cellular respiration - the includes the metabolic pathways which utilize oxygen and produce carbon dioxide, which will not be included in this unit. | |
| Structure of the respiratory system:
The upper respiratory division (See Figure 23.2) includes the nasal cavity and pharynx. Air enters the nasal cavity through the external nares and passes through the narrow channels (meatus) created between the nasal conchae. This exposes the air to the mucosa which warms and moistens the air as it passes through the entire system. The mucosa also removes particulates, dust, pollen, etc. and moves it toward the esophagus. In the nasal cavity the respiratory mucosa is partly ciliated and partly non ciliated and the cilia beat downward. [See olfactory epithelium] The upper portion of the nasal cavity has olfactory receptors whose nerves pass through the foramina in the cribriform plate to the olfactory bulb of the brain. The nasal cavity narrows at the internal nares before joining with the pharynx. The pharynx is divided into three portions: the nasopharynx, the oropharynx and the layngopharynx which connect to the nasal cavity, oral cavity and larynx respectively. The nasopharynx has the pharyngeal tonsils (lymph nodes) and the opening into the eustachian canal (a.k.a. pharyngotympanic tube or internal auditory canal). This canal allows equalization of pressure between the atmosphere and the middle ear. This is important when increasing or decreasing in altitude and when atmospheric pressure changes for any reason. Inability to equalize pressure can impair hearing as occurs when the mucosa is inflamed due to a respiratory infection. The lining continues to be ciliated into the nasopharynx and then changes to non-keratinized stratified squamous in the oropharynx and laryngopharynx where protection from ingested food is important. | |
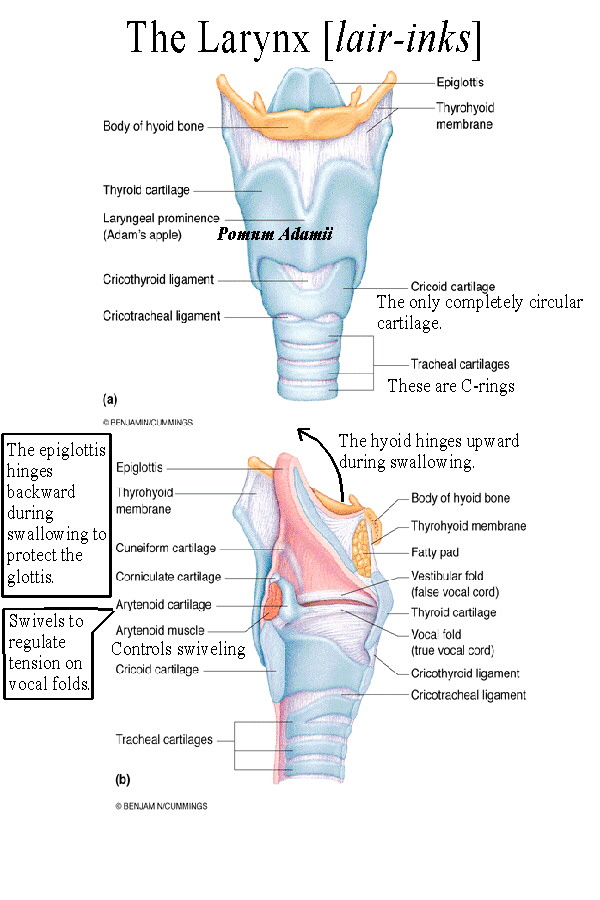 |
The larynx lies atop the respiratory tree and contains the voice box. Supporting the larynx is the hyoid bone which connects to the tongue above and to the thyroid cartilage below. When swallowing occurs, the hyoid bone, which does not articulate with any other bones, hinges upward and the larynx tilts backward. [See laryngeal histology] This causes the epiglottis to more effectively shield the glottis and prevent aspiration of food or liquid. The thyroid cartilage is the largest laryngeal cartilage and projects anteriorly as the "Adam's Apple". The thyroid cartilage is open posteriorly and the vocal folds (true vocal cords) run along its inside antero-posteriorly. Men tend to have deeper voices and more pronounced "Adam's Apples" due to the increased length of the thyroid cartilage and vocal fold. Tension on the vocal cords determines the pitch of the voice as air is pushed up between them. This tension in turn is controlled by cartilages attached to the back of the cords called the arytenoid cartilages. These cartilages swivel to change vocal cord tension. Vocal cord tension is controlled by arytenoid and other muscles [See laryngeal muscles] which contract or relax to increase (for high pitches) or decrease (for low pitches) tightness. Lying above the vocal folds are the vestibular folds (false vocal cords) which are narrow ridges protecting the vocal cords from aspirated materials. The glottis is the opening between the vocal cords and the passageway for air into the trachea. |
 |
The Conducting Zone - area
which conducts air into the lungs. No gas transport
occurs here. Also called the anatomical dead space.
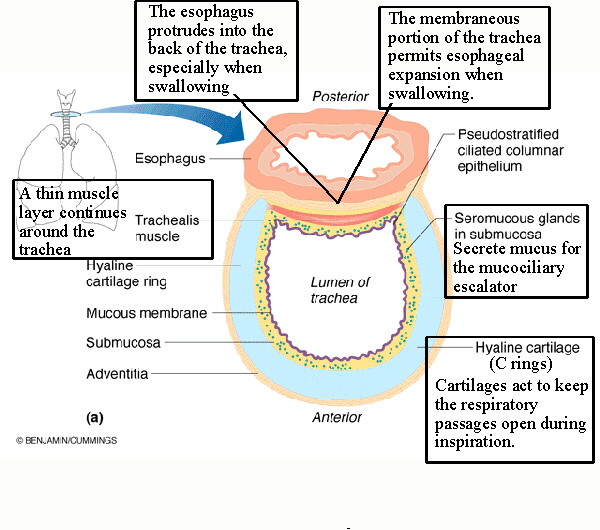
The trachea, or windpipe, is the beginning of the respiratory tree. It has cartilages shaped like Cs (or Us) with the open portion posterior. The trachea is lined (See tracheal lining) with pseudostratified ciliated columnar epithelium (p.c.c.e.) which continues into the bronchi. The cilia of this mucosa beat upward to carry particulates up and out into the esophagus. The submucosa of the trachea contains seromucous glands to produce an abundance of mucus for the mucociliary escalator, the mechanism by which the mucus is moved up and out of the respiratory tract. The membraneous portion of the trachea on the posterior side contains the majority of the trachealis muscle and is flexible to accommodate the esophagus nestled immediately behind the trachea. |
| The trachea branches into the primary
bronchi (See Figure 23.7 a) which lead to each
lung, then
branching to the secondary bronchi leading to the lobes (2 on the left, three on
the right), then the
tertiary
bronchi to the 18 broncho-pulmonary segments (8 on the left, 10 on the right).
Each
broncho-pulmonary segment includes a branch of the pulmonary artery, pulmonary vein, and a
tertiary
bronchus. These segments act as structural and functional units in the lung to maintain blood flow
and
gas transport. The cartilages change from C-rings to plate cartilages in the bronchi while the walls
get
thinner. PCCE continues.
Tertiary bronchi lead to the large bronchioles. Large bronchioles have little or no cartilage and the mucosa becomes simple ciliated columnar epithelium. Large bronchioles lead to the terminal bronchioles which have simple cuboidal epithelium. Goblet cells continue through the mucosa providing continued mucus secretion. | |
 |
The Respiratory Zone - this
area has thin simple squamous epithelial walls (the
respiratory bronchioles start out as simple cuboidal and soon become simple
squamous), no mucosa, and permits gas transport into
and out of the blood.
Respiratory bronchioles begin this zone, leading to alveolar ducts and then alveolar sacs. These sacs are a connected system of thin-walled chambers called alveoli. The alveoli increase the surface area for contact with blood vessels.(See Figure 23.8, modified) The structure of the lungs is composed of these spongy-appearing alveolar sacs, together with bronchial passageways and blood vessels. See the following for highly magnified views of alveoli and associated cells and tissues: [alveoli 1][alveoli 2] Look for the numerous capillaries in these closeups. They are indicated by the red blood cells inside them. |
 |
Alveolar capillaries are more numerous and densely arranged than in most any other organ. They form an area of contact with the alveolar walls of (depending on which source you read) from 50 to 400 sq. meters. (See Figure from class) Upon this area, at rest, approximately a liter and a half of blood is distributed. (This amount increases with exercise). A dense network of capillaries surrounds each alveolus to produce the respiratory membrane (See Figure 23.9) consisting of the capillary wall (endothelium), the alveolar wall (simple squamous epithelium), and the basement membranes of each. These basement membranes are composed of a basal lamina with collagen fibers and a small amount of elastic tissue. The elastic tissue gives the lungs their elasticity or recoil. The structure of the respiratory membrane permits gas transport while preventing much water from entering the alveoli from the capillaries. Since interstitial fluid in the lungs would be disastrous to respiratory function, any fluid which does enter is removed by the lymph system. All the moisture in the alveolar sacs is the result of evaporation from the mucosal lining of the respiratory passages above. In addition to the Type I alveolar cells (the simple squamous cells of the alveolar wall) there are Type II alveolar cells present which secrete surfactant. Surfactant acts to break the surface tension (cohesiveness) of water which would cause collapse of the alveoli during expiration. This happens in premature infants whose respiratory systems have not matured, a condition called Infant Respiratory Distress Syndrome (IRDS) (DEF) or Hyaline Membrane Disease . Macrophages are also present which phagocytize bacteria that make it into the alveoli. |
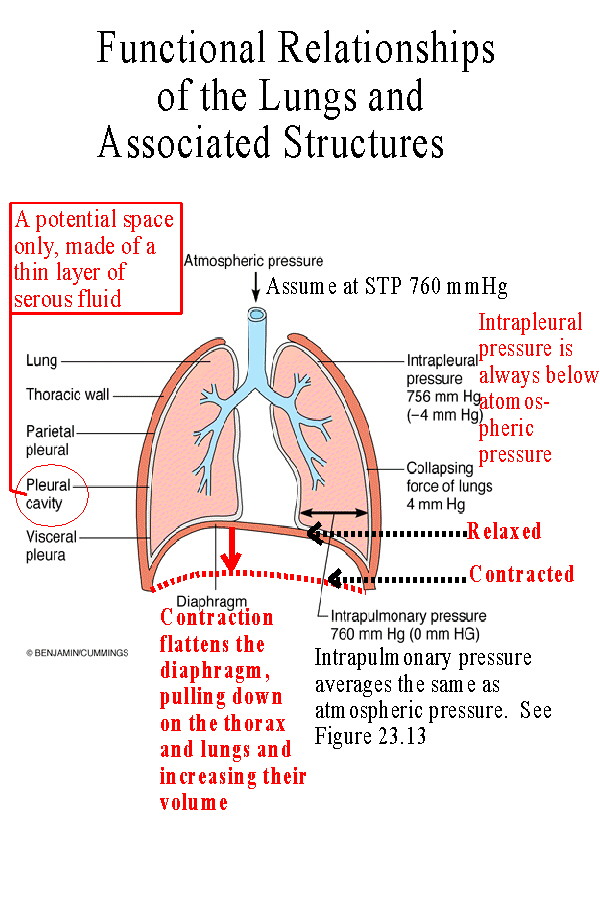 |
(See Figure 23.10 and 23.11)The lungs are surrounded by a double layered pleural membrane or sack. The inner layer is called the visceral pleura and is attached to the surface of each lung. The outer layer is the parietal pleura and is attached to the lining of the thorax and the mediastinum. It adheres to the inner wall of the ribcage and to the upper surface of the diaphragm. The space between these layers, sometimes called the pleural space or cavity, is really only a potential space, normally just the layer of serous fluid secreted by the membranes. The serous fluid binds the two layers together so that if the parietal layer is expanded by movement of the ribcage or diaphragm, so is the visceral pleura and so are the lungs. |
| Ventilation is composed of two parts: inspiration and expiration. Each of these can be described as being either quiet, the process at rest, or forced, the process when active such as when exercising. Boyle's Law states that the volume and pressure of a gas are inversely proportional. If the volume of the gas increases, its pressure will decrease. If the volume decreases, its pressure will increase. The movement of air in ventilation occurs as a result of the pressure gradient produced when the volume of the lungs increases or decreases. The following table describes the events which produce this pressure gradient: | |
| Quiet
inspiration:
The diaphragm contracts, this causes an increase in volume of the thorax and the lungs, which causes a decrease in pressure of the thorax and lungs, which causes air to enter the lungs, moving down its pressure gradient. Air moves into the lungs to fill the partial vacuum created by the increase in volume. |
Forced
inspiration:
Other muscles aid in the increase in thoracic and lung volumes. The scalenes - pull up on the first and second ribs. The sternocleidomastoid muscles pull up on the clavicle and sternum. The pectoralis minor pulls forward on the ribs. The external intercostals are especially important because they spread the ribs apart, thus increasing thoracic volume. (See Figure 23.12) It's these muscles whose contraction produces the "costal breathing" during rapid respirations. |
| Quiet expiration:
The diaphragm relaxes. The elasticity of the muscle tissue and of the lung stroma causes recoil which returns the lungs to their volume before inspiration. The reduced volume causes the pressure in the lungs to increase thus causing air to leave the lungs due to the pressure gradient. |
Forced Expiration:
The following muscles aid in reducing the volume of the thorax and lungs: The internal intercostals - these compress the ribs together (See Figure 23.12). The abdominus rectus and abdominal obliques: internal obliques, external obliques- these muscles push the diaphragm up by compressing the abdomen. |
| Respiratory output is determined by the
minute volume, calculated by multiplying the
respiratory rate time the tidal volume.
Minute Volume = Rate (breaths per minute) X Tidal Volume (ml/breath) Rate of respiration at rest varies from about 12 to 15 bpm. Tidal volume averages 500 ml (See Figure 23.15 or Lung Volumes Table below) Assuming a rate of 12 breaths per minute and a tidal volume of 500, the restful minute volume is 6000 ml. Rates can, with strenuous exercise, increase to 30 to 40 bpm and volumes can increase to around half the vital capacity. Not all of this air ventilates the alveoli, even under maximal conditions. The conducting zone volume is about 150 ml and of each breath this amount does not extend into the respiratory zone. The Alveolar Ventilation Rate, AVR, is the volume per minute ventilating the alveoli and is calculated by multiplying the rate times the (tidal volume-less the conducting zone volume). AVR = Rate X (Tidal Volume - 150 ml) For a calculation using the same restful rate and volume as above this yields 4200 ml. Since each breath sacrifices 150 ml to the conducting zone, more alveolar ventilation occurs when the volume is increased rather than the rate. | |
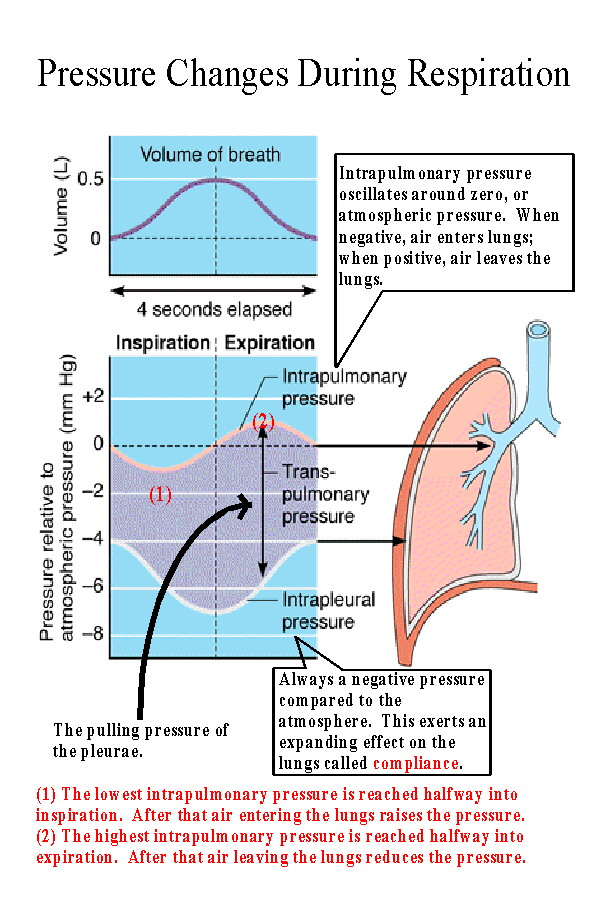 |
During inspiration the pressure inside the lungs (the
intrapulmonary pressure) decreases to -1 to -3 mmHg compared
to the atmosphere. The variation is related to the forcefulness and
depth of inspiration. During expiration the intrapulmonary pressure
increases to +1 to +3 mmHg compared to the atmosphere. The
pressure oscillates around zero or atmospheric pressure.
The intrapleural pressure is always negative compared to the atmosphere. This is necessary in order to exert a pulling action on the lungs. The pressure varies from about -4 mmHg at the end of expiration, to -8 mmHg and the end of inspiration. |
| The tendency of the lungs to expand, called
compliance or distensibility, is due to
the pulling action exerted by the pleural membranes. Expansion is also facilitated by
the action of surfactant in preventing the collapse of the alveoli.
The opposite tendency is called elasticity or recoil, and is the process by which the lungs return to their original or resting volume. Recoil is due to the elastic stroma of the lungs and the series elastic elements of the respiratory muscles, particularly the diaphragm. NOTE: Click on names for comprehensive review of respiratory conditions or on (def) for expanded definition. Also see the Respiratory Pathology Images Conditions which interfere with compliance or elasticity are called restrictive disorders. Examples are emphysema (def), which increases compliance and decreases elasticity, and fibrosis (def), which reduces both. In emphysema the buildup of toxins from cigarette smoke and resulting mucus production leads to destruction of the alveolar and capillary walls and fibrosis of the tissue. This produces large thick-walled chambers replacing the normal small thin-walled alveoli. It results in a larger volume in the lungs but impaired gas transport and reduced ability to expire the trapped air. Many emphysema sufferers have the characteristic "barrel chest" as a result. Carbon dioxide tends to increase in alveolar air and in the blood, in some individuals interfering with normal respiratory stimuli and responses. Pulmonary or cystic fibrosis (def) produces thickened mucus secretions which block the airways and scar tissue often develops in place of the normal elastic stroma. This is both restrictive and obstructive in its effects. Restrictive disorders reduce the volume which can be ventilated as seen in a reduced vital capacity. (See below and Figure 23.15) In contrast, obstructive disorders such as bronchitis (def) and asthma (def) reduce the size of the bronchial passages thus interfering with the airflow. [See COPD below] In bronchitis an inflammation results from respiratory infection or irritation from smoke or pollution. Asthma is like an allergic reaction in which inflammation also occurs together with a constriction of the bronchial passages. Purely obstructive disorders do not technically reduce the volume which can be ventilated but they reduce the rate of ventilation by increasing the resistance to airflow. They will show a normal vital capacity but a reduced FEV1, (Forced Expiratory Volume) the percentage of the vital capacity which can be expelled in the first second. Normal FEV1 is 75% or greater. Many times disorders are both obstructive and restrictive. For example: obstructive emphysema - in addition to the enlarging and thickening of alveolar walls many of the passageways collapse producing obstructive effects as well. COPD Chronic Obstructive Pulmonary Disease - a combination of emphysema, bronchitis and asthma, has components of each. Other respiratory conditions: SIDS - Sudden Infant Death Syndrome | |
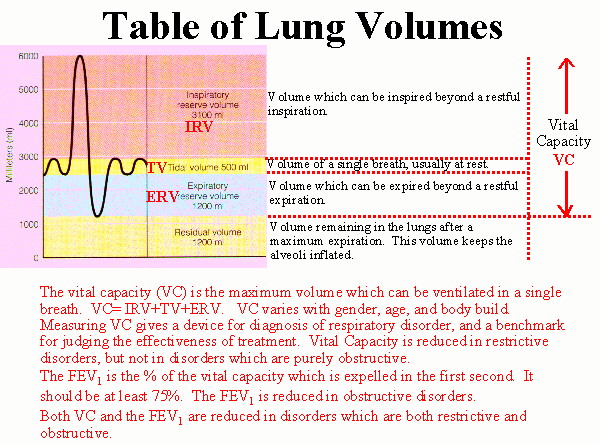 |
Respiratory volumes
Tidal Volume TV: Volume of a single breath, usually at rest. Inspiratory Reserve Volume IRV: Volume which can be inspired beyond a restful inspiration. Expiratory Reserve Volume ERV: Volume which can be expired beyond a restful expiration. Residual Volume RV: Volume remaining in the lungs after a maximum expiration. This volume keeps the alveoli inflated. Vital Capacity: The vital capacity (VC) is the maximum volume which can be ventilated in a single breath. VC= IRV+TV+ERV. VC varies with gender, age, and body build. Measuring VC gives a device for diagnosis of respiratory disorder, and a benchmark for judging the effectiveness of treatment. Vital Capacity is reduced in restrictive disorders, but not in disorders which are purely obstructive. The FEV1 is the % of the vital capacity which is expelled in the first second. It should be at least 75%. The FEV1 is reduced in obstructive disorders. Both VC and the FEV1 are reduced in disorders which are both restrictive and obstructive. (See above) |
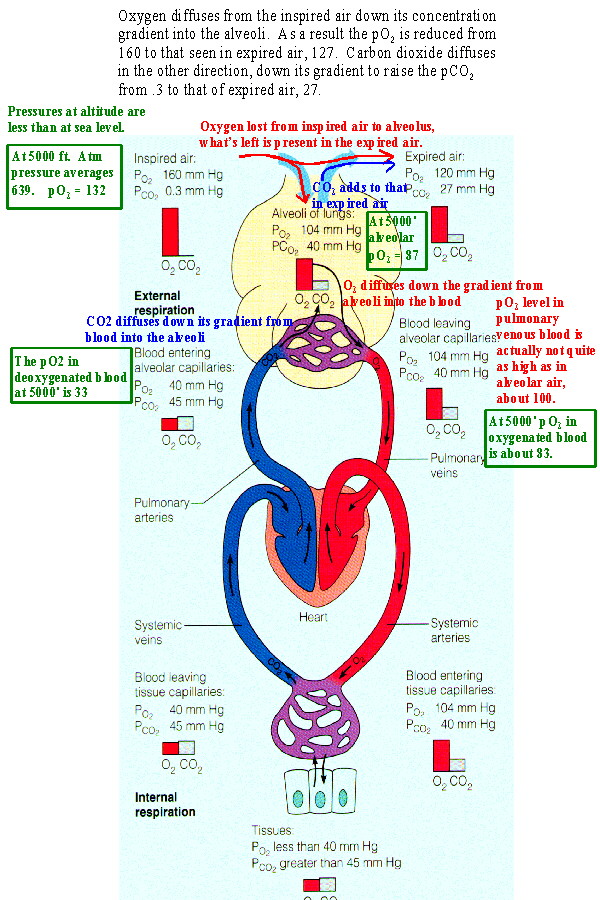 |
Movement of the respiratory gases is due to diffusion. Diffusion
results from a concentration gradient which is expressed for gases as
the difference in partial pressures.
Dalton's Law of Partial Pressures states that The partial pressure of a gas in a mixture is calculated by multiplying the fraction occupied by the gas times the total pressure of the mixture. The mixture of gases under consideration is the air and Table 23.4 shows the partial pressures of its gases at STP (standard temperature and pressure at sea level). Oxygen is present at nearly 21% of ambient air. Multiplying .21 times 760 mmHg (standard pressure at sea level) yields a pO2 of about 160. Carbon dioxide is .04% of air and its partial pressure, pCO2, is .3. As Figure 23.16 shows there is a partial pressure gradient between inspired air and the alveolus with alveolar air having a pO2 of 104 and a pCO2 of 40. So oxygen diffuses into the alveoli from inspired air and carbon dioxide diffuses from the alveoli into air which will be expired. This causes the levels of oxygen and carbon dioxide to be intermediate in expired air when compared to inspired air and alveolar air. Some oxygen has been lost to the alveolus, lowering its level to 120, carbon dioxide has been gained from the alveolus raising its level to 27. Likewise a concentration gradient causes oxygen to diffuse into the blood from the alveoli and carbon dioxide to leave the blood. This produces the levels seen in oxygenated blood in the body. When this blood reaches the systemic tissues the reverse process occurs restoring levels seen in deoxygenated blood. |
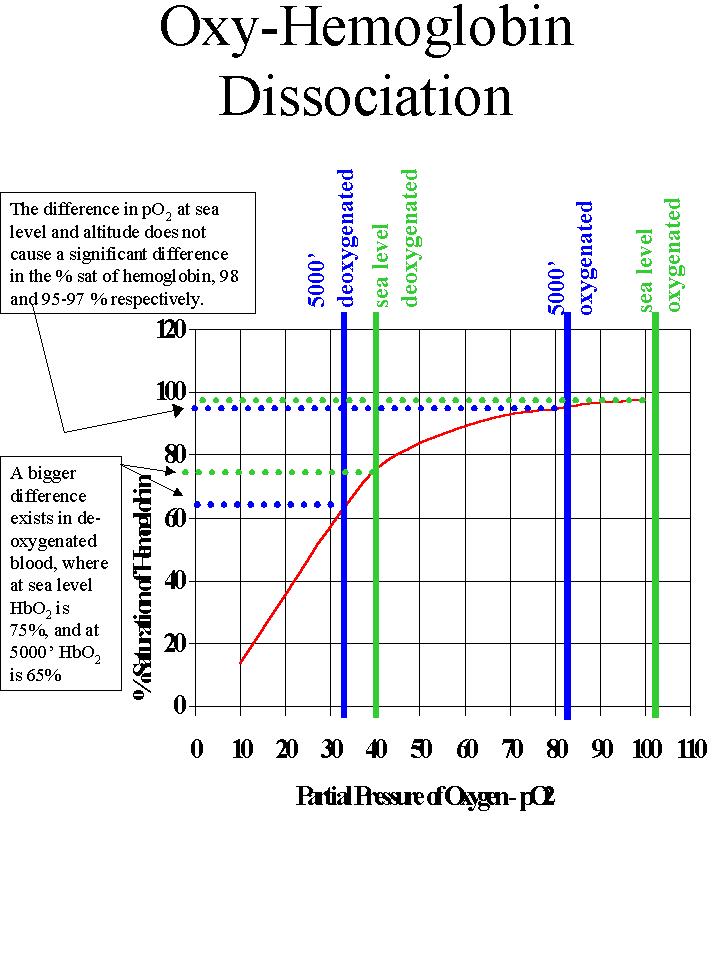 |
The effect of pO2 on hemoglobin
saturation - the Oxyhemoglobin
Dissociation Curve. Figure 23.19
This curve illustrates the % saturation of hemoglobin at different levels of pO2 . Because the curve is flat on top, changes in pO2 in this region do not result in significant differences in % saturation of hemoglobin. At sea level with a pO2 in oxygenated blood of 100+ (as seen earlier) the % saturation of hemoglobin is 98%. At 5000' the pO2 in oxygenated blood has decreased to about 83, but the saturation of hemoglobin has gone down only a few % to 95-97. Deoxygenated blood is in the steeper portion of the curve. At sea level the pO2 in deoxygenated blood is 40. This results in saturation of hemoglobin of about 75%. This means that about 1/4 of the oxygen carried by hemoglobin has been released to the tissues. The remaining oxygen on hemoglobin represents a significant reservoir which can be delivered to the tissues during hypoxic stress. At 5000' the pO2 is about 33. This results in a saturation of hemoglobin of 65%. More oxygen has been unloaded (dissociated) from hemoglobin at altitude. The result of lowered pO2, whether as a result of altitude or hypoxic stress is that more oxygen is unloaded to the tissues from the oxygen reservoir. |
 |
The effect of pH on the Oxyhemoglobin Dissociation Curve.
(Figure 23.20).
The effect of lowered pH steepen the curve resulting in greater dissociation or unloading of oxygen from hemoglobin. Higher pH flattens the curve resulting is reduced dissociation or unloading of oxygen from hemoglobin. This is due to a coupling of chemical reactions in which the reduction in pH which occurs as the blood picks up CO2 in the systemic tissues enhances the unloading of oxygen from hemoglobin. Likewise the increase in pH accompanying release of CO2 from the blood to alveolar air causes hemoglobin to load up with oxygen. |
| The Bohr Effect describes the result of increasing CO2 in causing more oxygen unloading from hemoglobin. [See Bohr Effect] It results from two circumstances: 1) the effect of lowering pH as described above, and 2) the effect of carbaminohemoglobin in stimulating oxygen unloading. The corollary to the Bohr Effect is called the Haldane Effect. This effect states that a reduction of oxygen on the hemoglobin molecule allows for more CO2 transport as carbaminohemoglobin. | |
| Control of respiration: (See Figures 23.22,
23.23)
The respiratory center is located in the medulla of the brainstem, with contributions from the pons. Within the medulla are the DRG (dorsal respiratory group) and the VRG (ventral respiratory group). The DRG sends stimuli to the muscles of inspiration: the diaphragm, external intercostals, and other. The VRG sends stimuli to the muscles of expiration: the internal intercostals and abdominal muscles. Since these muscles act only in forced expiration the VRG is only active then, while the DRG acts in both quiet and forced respiration. These muscles are all skeletal muscles and the motor control of respiration, whether quiet or forced, is a voluntary function. The autonomic nervous system does not control the muscles of respiration. The medulla controls the rhythmicity of respiration. The pons sends stimuli to the medulla to regulate the rate and depth. The pneumotaxic center increases the rate by shortening inspirations. The apneustic center (its function is established but its location is not) increases the depth and reduces the rate by prolonging inspirations. Inputs to these centers come from several locations: 1) peripheral chemoreceptors - located in the aortic sinus and carotid sinus, respond to increased carbon dioxide and decreased pH (which are related as discussed earlier). Although the peripheral chemoreceptors include receptors to oxygen, oxygen levels are secondary stimuli at best, normally not important in triggering increased respiration. For healthy people the levels of oxygen would never decrease to the point of stimulating respiration. However, this can apparently happen in certain disorders such as emphysema, in a condition known as hypoxic drive. This happens because CO2 levels become permanently elevated and the CO2 receptors cease responding. This is somewhat controversial, but some believe that administering oxygen can actually suppress respiration in someone with hypoxic drive. 2) central chemoreceptors - located in the medulla. Their primary stimuli are decreased pH and increased carbon dioxide. pH is especially important since carbon dioxide does not readily diffuse into brain tissue but affects it through its impact on hydrogen ions. 3) muscle contraction - when you exercise there is a direct stimulus to the respiratory center from active muscles and joint receptors. This causes increased respiration before blood chemistry actually changes enough to demand it. 4) higher brain centers - from the voluntary motor center for voluntary control over respiration, and from the hypothalamus for control in response to emotional stimuli and body temperature. 5) vagal afferents - the only autonomic part of respiration, these send stimuli from stretch receptors in the lungs and from irritant receptors. The stretch receptors allow a reflex which prevents overinflation called the Hering-Breuer Reflex, much as a skeletal muscle reflexively changes from one action to another. The irritant receptors act to produce coughing, hiccups, etc. | |
The following table summarizes the responses to various stimuli on the respiratory system:
| Stimulus | Response |
| primary: increased blood CO2, decreased pH,
secondary: decreased O2 These stimuli can come from respiratory or non-respiratory causes |
hyperventilation
(increase in rate and/or volume) |
| decreasing CO2, increasing pH, increased O2 | hypoventilation |
| voluntary (whether intentional or not)
hyperventilation |
reduced CO2 in blood, increased pH
reduced stimulus for respiration |
| voluntary hypoventilation | increased CO2, decreased pH
increased stimulus for respiration. |
NEXT: Digestive System
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}