Physiologists refer to receptors as selective transducers. They are called transducers because they 'convert' the energy contained in the stimulus into another form of energy, specifically into some sort of membrane potential. Receptors are termed selective because each type of receptor is highly specific (selective) with respect to the type of stimulus it responds to. Thus (in the absence of LSD or similar compounds), the eye doesn't transduce sound or temperature, the ear doesn't transduce light or temperature, and so on.
Although there are many different types of receptors, all receptors go about the actual transduction of stimulus into response in the same basic fashion. In brief, the energy contained in the stimulus is used by the receptor to change the conductance of its membrane for one or more ions. Of course, the conductance change(s) will cause the receptor's membrane potential (Vm) to change. Depending on the nature of the change in Vm (i.e., depolarization, or hyperpolarization), the frequency of action potentials sent to the brain along the receptor's sensory neuron will increase or decrease in response to the change in Vm. This is illustrated in the following block diagram:
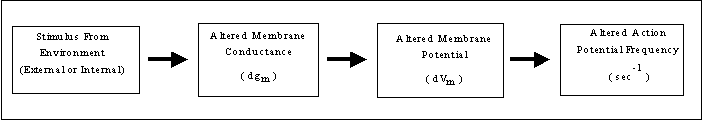
Now, let's discuss the steps in the diagram with reference to a specific receptor that you're somewhat familiar with as a result of the Senses lab you did earlier this semester: the Pacinian Corpuscle (cf. Fig. 42.9 in your text). The Pacinian Corpuscle is one of a variety of receptors located in your skin, and is and ideal model receptor to use for discussion of how receptors transduce stimuli. Pacinian Corpuscles are examples of what we call mechanoreceptors because what they respond to is pressure applied to the skin's surface. Other mechanoreceptors include stretch receptors in your tendons and the receptors in your ear that respond to sound.
The Pacinian Corpuscle is an excellent model for understanding how receptors work. It consists of two components: the tip of a sensory neuron that conducts action potentials towards the brain and a number of layers of 'membrane', called lamellae, wrapped around the neuron tip. In cross section, the Pacinian corpuscle looks like a little onion:
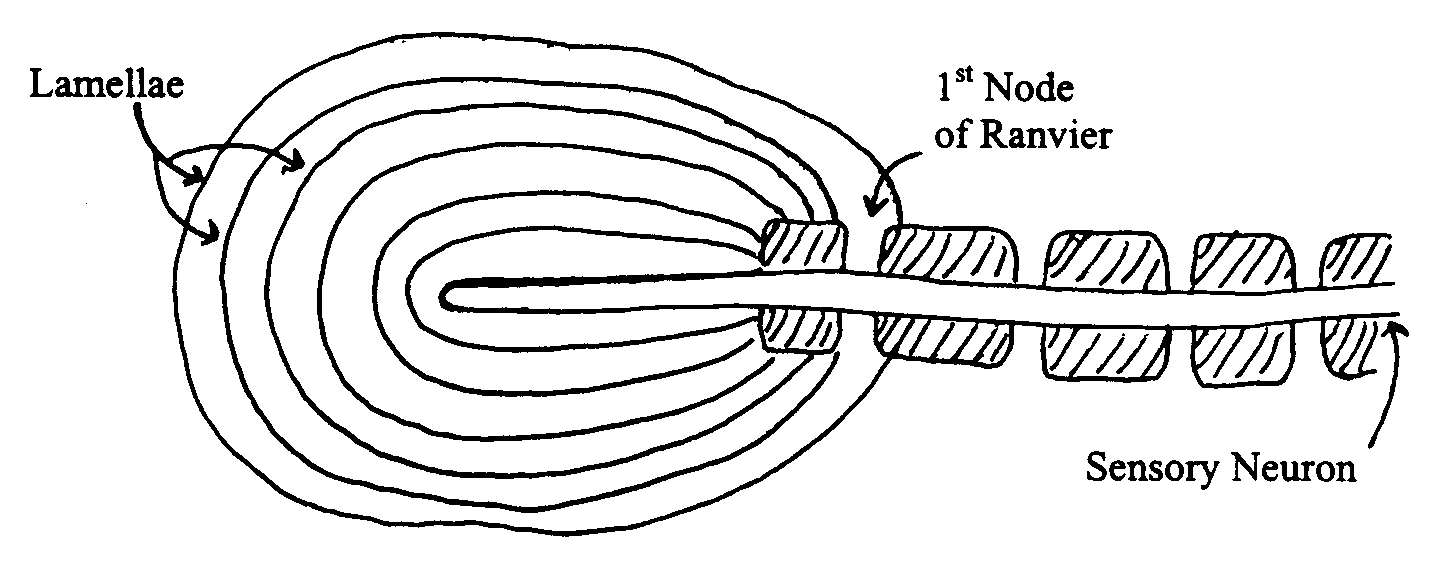
When you press on a Pacinian Corpuscle, you deform the lamellae and cause them to press on the tip of the sensory neuron. That, in turn, physically deforms the neuron's plasma membrane and makes it 'leaky' to sodium ions (i.e., the deformation increases gNa+). With your background in membrane electrical phenomena, you of course recognize that this will tend to depolarize the membrane at the site of the deformation. The tip of the sensory neuron, however, is like the dendritic zone of a neuron in that it lacks voltage-gated Na+ channels and therefore cannot generate or propagate action potentials, so you can think of this depolarization as a large-amplitude EPSP (dVm = 20-50 mV, depending on how strong the pressure is).
The first part on the sensory neuron that does have the voltage-gated Na+ channels required for action potential generation is the 1st Node of Ranvier. In a way similar to the way an EPSP depolarizes the Initial Segment of an axon to threshold, the depolarization at the tip of the neuron depolarizes the 1st Node of Ranvier to threshold and causes it to start generating action potentials which propagate toward the brain. So, in a sense the 1st Node of Ranvier is serving as an Initial Segment.
As long as the tip of the neuron is depolarized, it will continue to depolarize the 1st Node of Ranvier, which will respond by generating action potentials as long as the depolarization lasts. This is because as soon as the 1st Node of Ranvier recovers from generating an action potential, the depolarized neuron tip depolarizes the 1st Node of Ranvier back to its threshold. Then, as soon as the depolarized 1st Node of Ranvier 'gets out of' its absolute refractory period (which is quite short as refractory periods go) it generates another action potential. This continues as long as the neuron tip remains depolarized.
Because the initial depolarization of the neuron tip tends to cause the 1st node of Ranvier to generate action potentials, the depolarization was originally termed a generator potential by the investigator who discovered it. This term was soon generalized to the dVm exhibited by all receptors. This was unfortunate, because many receptors actually hyperpolarize when stimulated, which of course would not tend to generate action potentials. For that reason, many physiologists now use the term receptor potential to refer to the initial dVm of a receptor when it's stimulated. I prefer that latter term, but either is acceptable.
Encoding The Strength Of The Stimulus
It's generally not enough for the receptor simply to inform the brain that it's being stimulated; information about the strength of the stimulus is also desirable. Because action potentials 'obey' the All-or-None law (p. 912 in your text), the strength of the stimulus cannot be encoded by varying the amplitude of the action potentials (= amplitude modulation). Instead, the receptor varies the frequency of action potentials sent to the brain. This encoding method is termed frequency modulation, and it the only way that receptors can inform the brain about the strength of the stimulus impinging on them.
In the case of the Pacinian Corpuscle, frequency modulation is possible because a stronger pressure produces a larger receptor potential (i.e., a larger depolarization) which has a stronger depolarizing effect on the 1st Node of Ranvier and depolarizes it to threshold more rapidly. Thus, it is the frequency of action potentials being sent to the brain by the Pacinian Corpuscle that 'informs' the brain about the strength of the stimulus (i.e., the amount of pressure being applied to the corpuscle).
A graph of action potential frequency vs. stimulus intensity looks like this:
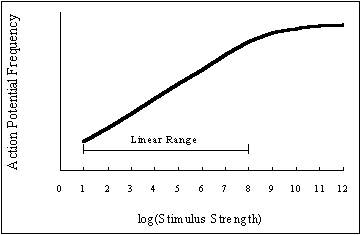
Now, look carefully at the graph. Although it may be a little tricky to tell depending on your monitor's performance and settings, the response of the receptor to changes in log(Stimulus Strength) is actually quite linear over a range of 8 magnitudes of Stimulus Strength. And, the frequency of action potentials generated by the receptor actually continues to increase until it 'plateaus' as log(Stimulus Strength ) exceeds a value of roughly 11. What this means is that the receptor can use variation in action potential frequency to encode Stimulus Strengths that vary by nearly 11 orders of magnitude. Consequently, the brain can determine the strength of the stimulus that's being applied to the receptor from the frequency of action potentials arriving along the sensory neuron.
Although there are differences in the details of function in various receptors, you now know the basics of how all receptors transduce the information contained in a stimulus and forward it to the brain.